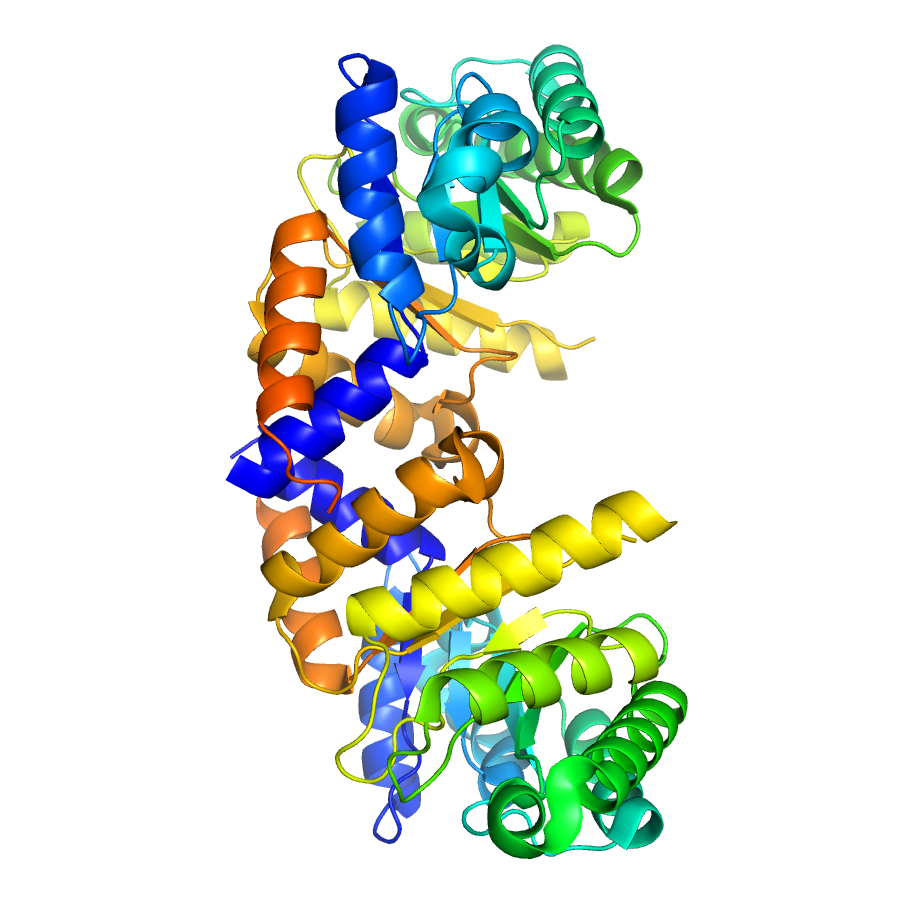
Structure of IDP90300
1.37 Angstrom resolution crystal structure of apo-form of a putative deoxyribose-phosphate aldolase from Toxoplasma gondii ME49
Edit deposit information
- CSGID target
- IDP90300
- PDB Id
- 4EIV (NCBI MMDB)
- Authors
- A.S.Halavaty,J.Ruan,G.Minasov,L.Shuvalova,A.Ueno,M.Igarashi,H.Ngo,W.F.Anderson,Center For Structural Genomics Of Infectious Diseases (Csgid)
- Responsible person
- Andrei Halavaty
- Responsible lab
- Northwestern University
- Deposition Date
- Apr 05, 2012
- Release Date
- May 23, 2012
Annotation
- Description
- Toxoplasma gondii deoxyribose phosphate aldolase-like (TgDPA) gene is expressed predominantly in bradyzoites, indicating it could be important in the tachyzoite-to-bradyzoite development or the maintenance of cyst structure. Yeast two-hybrid screening, Co-immunoprecipitation and a GST pull-down assays have demonstrated that TgDPA interacts with the actin depolymerizing factor (TgADF) in bradyzoite stage of the parasite and enhances the actin depolymerizing activity of TgADF. TgDPA lost its deoxyribose phosphate aldolase enzymatic function due to the substitution of the key residue, K173, to a glutamine residue at the activity site, therefore, this potential drug target protein seems to play a novel, important role in maintaining the T. gondii bradyzoites stage by affecting the parasite’s actin turnover. We have recently determined the second 3D structure of TgDPA (PDB code 4EIV) by molecular replacement and refined it to 1.37 Å. The protein is considered to be in its apo form in the new P212121 space group, whereas the protein in the first structure (PDB code 3QYQ; P1 space group) was suggested to be in its product/substrate-bound form. The latter assumption was made based on the presence of cryo-protectant sucrose molecule(s) in the vicinity of the active site. Since TgDPA is not a deoxyribose phosphate aldolase and may have a new role(s) (see above), our current TgDPA structure represents a new crystal form. Similar to the first structure, TgDPA exhibits (α8/β8)-TIM-barrel topology and chains in both structures have RMSD values between 0.7 and 1.1 Å in the position of their C-alpha atoms. In the current 4EIV structure, the C-terminal 10 residues of the 40-Å long helix α6 observed in the 3QYQ structure are disordered because of the crystal packing. Although the TIM-barrel core remained rigid in the two crystal forms, forced disorder/flexibility of the helix α6 may be a key structural feature in binding TgADF. Additionally, residues 147 through 153 and residues 175 through 189 that form surface loops at the C-termini of the TIM-barrel structure exhibit different conformations in both structures. These structural changes may be a result of a new crystal environment and/or the absence of ligands such as sucrose seen in the 3QYQ structure. Moreover, small movements at the dimerization interface as well in some solvent (crystal packing)-exposed areas of the protein may be also related to the new crystal form.
- Functional assignment
- Lyase
Ligands
| Ligand code | Name | Ligand type |
|---|---|---|
| CL | chloride | crystallization |
| 175 | 3,5-dihydro-5-methylidene-4h-imidazol-4-on |
Structure information
Unit cell parameters
- Space Group
- P 21 21 21
- Unit Cell
-
a=71.98Å, b=73.28Å, c=96.38Å
α=90.00, β=90.00, γ=90.00 - Solvent content
- Matthews coefficient
- Resolution range
- 29.42-1.37Å (1.41-1.37Å)
- Rall(%)
- 15.3
- Rwork(%)
- 15.2 (20.8)
- Rfree(%)
- 18.5 (23.4)
- Num. observed reflections
- 106887 (7879)
- Num. Rfree reflections
- 5344 (400)
- Completeness(%)
- 99.4 (100.0)
- Num Atoms
- 4208
- Num Waters
- 688
- Num Hetatoms
- 732
- Model mean isotropic B factor
- 34.300Å2
- RMSD bond length
- 0.010Å
- RMSD bond angle
- 1.428°
- Filename uploaded
- 4EIV.pdb (uploaded on Jun 04, 2015 3:47 PM)
- Inserted
- May 04, 2012