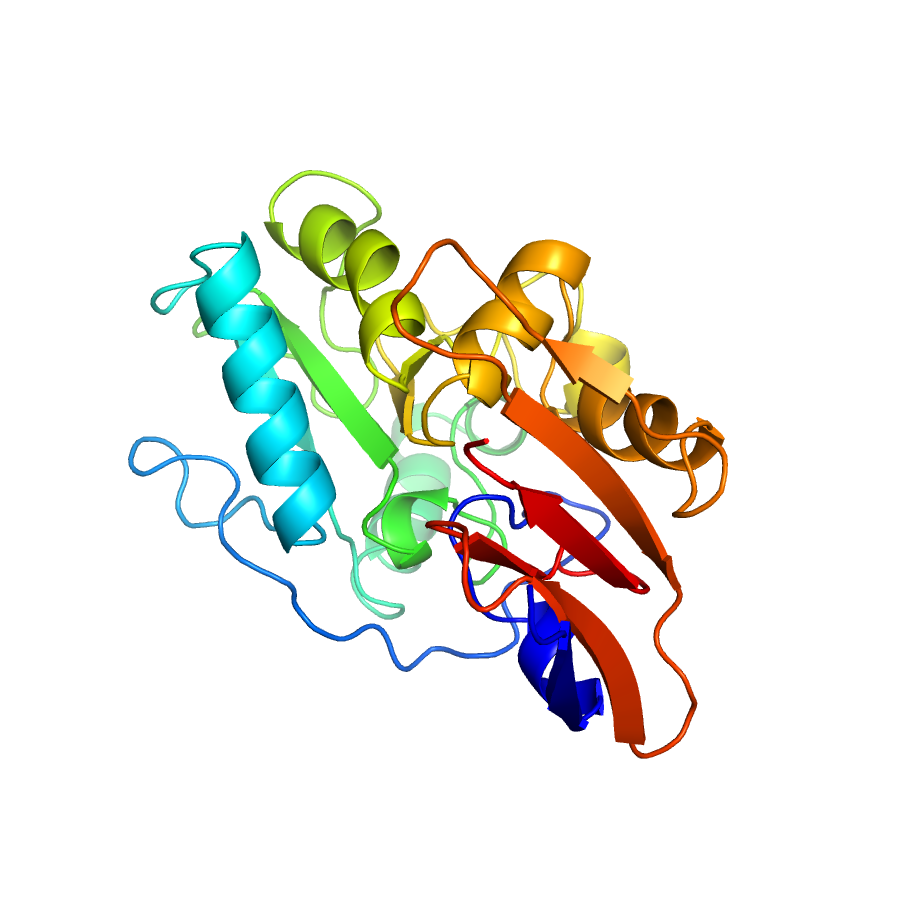
Structure of IDP91774
Crystal Structure of Vancomycin Resistance D,D-dipeptidase VanXYg
Edit deposit information
- CSGID target
- IDP91774
- PDB Id
- 4F78 (NCBI MMDB)
- Authors
- P.J.Stogios,Z.Wawrzak,E.Evdokimova,G.Minasov,O.Egorova,R.Di Leo,M.Kudritska,V.Yim,D.Meziane-Cherif,P.Courvalin,A.Savchenko,W.F.Anderson,Center For Structural Genomics Of Infectious Diseases (Csgid)
- Responsible person
- Peter Stogios
- Responsible lab
- University of Calgary
- Deposition Date
- May 15, 2012
- Release Date
- May 23, 2012
Annotation
- Description
- Vancomycin resistance in Gram-positive bacteria is due to production of cell-wall precursors ending in d-Ala-d-Lac or d-Ala-d-Ser, to which vancomycin exhibits low binding affinities, and to the elimination of the high-affinity precursors ending in d-Ala-d-Ala. Depletion of the susceptible high-affinity precursors is catalyzed by the zinc-dependent d,d-peptidases VanX and VanY acting on dipeptide (d-Ala-d-Ala) or pentapeptide (UDP-MurNac-l-Ala-d-Glu-l-Lys-d-Ala-d-Ala), respectively. Some of the vancomycin resistance operons encode VanXY d,d-carboxypeptidase, which hydrolyzes both di- and pentapeptide. The molecular basis for the diverse specificity of Van d,d-peptidases remains unknown. We present the crystal structures of VanXYC and VanXYG in apo and transition state analog-bound forms and of VanXYC in complex with the d-Ala-d-Ala substrate and d-Ala product. Structural and biochemical analysis identified the molecular determinants of VanXY dual specificity. VanXY residues 110-115 form a mobile cap over the catalytic site, whose flexibility is involved in the switch between di- and pentapeptide hydrolysis. Structure-based alignment of the Van d,d-peptidases showed that VanY enzymes lack this element, which promotes binding of the penta- rather than that of the dipeptide. The structures also highlight the molecular basis for selection of d-Ala-ending precursors over the modified resistance targets. These results illustrate the remarkable adaptability of the d,d-peptidase fold in response to antibiotic pressure via evolution of specific structural elements that confer hydrolytic activity against vancomycin-susceptible peptidoglycan precursors.
- Functional assignment
- antibiotic resistance
Ligands
| Ligand code | Name | Ligand type |
|---|---|---|
| EDO | ethylene diol | crystallization |
| SO4 | sulfate | crystallization |
| CL | chloride | crystallization |
| ZN | zinc | biological |
| MSE | modified residue | |
| 175 | 3,5-dihydro-5-methylidene-4h-imidazol-4-on |
Structure information
Unit cell parameters
- Space Group
- P 1 21 1
- Unit Cell
-
a=38.91Å, b=43.96Å, c=80.20Å
α=90.00, β=99.19, γ=90.00 - Solvent content
- Matthews coefficient
- Resolution range
- 39.58-1.95Å (2.00-1.95Å)
- Rall(%)
- 15.5
- Rwork(%)
- 15.4 (21.7)
- Rfree(%)
- 18.6 (27.8)
- Num. observed reflections
- 20683 (2747)
- Num. Rfree reflections
- 1032 (123)
- Completeness(%)
- 98.3 (98.0)
- Num Atoms
- 2055
- Num Waters
- 187
- Num Hetatoms
- 296
- Model mean isotropic B factor
- 26.700Å2
- RMSD bond length
- 0.016Å
- RMSD bond angle
- 1.494°
- RMSD dihedral angle
- 14.268°
- Filename uploaded
- 4F78.pdb (uploaded on Apr 23, 2014 10:01 AM)
- Inserted
- May 15, 2012